
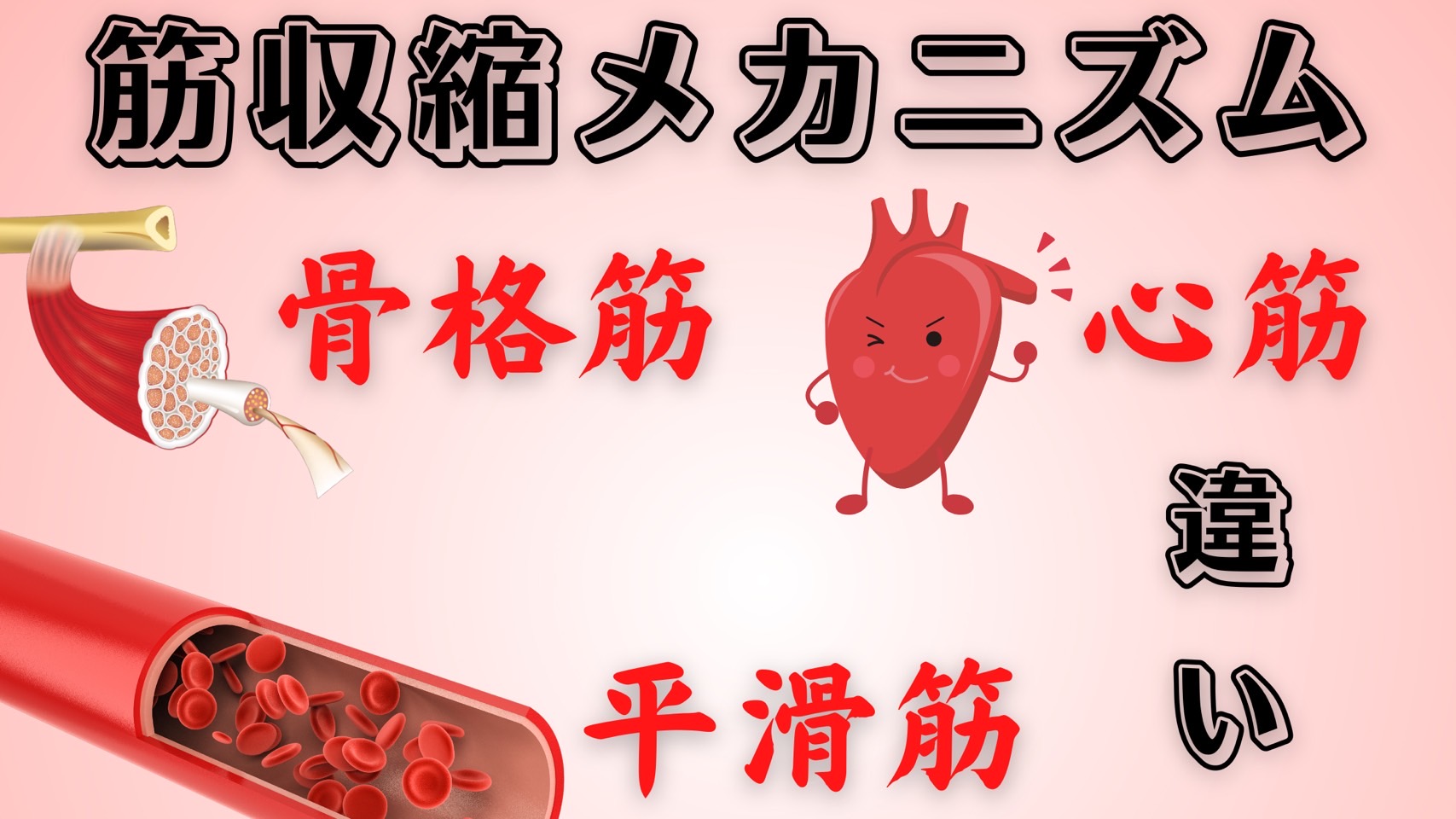
骨格筋の筋収縮メカニズム

①中枢からの骨格筋収縮の指令は活動電位により電気的信号として運動神経線維を伝わってきて運動神経終末までやってくる。運動神経線維は自律神経のように中枢から支配器の間で神経節でニューロン交代(乗り換え)をすることがないので、節前線維と節後線維という区別がなく1本である。
②活動電位が伝わるとシナプス前膜が脱分極して電位依存性カルシウムイオンチャネルが開く。
③カルシウムイオンが運動神経終末に流入してきて運動神経細胞内のカルシウムイオン濃度が上がると、シナプス小胞がシナプス前膜と膜融合してアセチルコリンをシナプス間隙に開口分泌(exocytosis)する。ここで活動電位という電気的信号がアセチルコリンという化学的信号に変わることになる。
④放出されたアセチルコリンは運動終板上にあるニコチン性アセチルコリン受容体(Nm受容体)に結合する。
⑤ニコチン性アセチルコリン受容体(Nm受容体)にアセチルコリンが結合するとナトリウムイオンが流入して運動終板が脱分極する。ここでアセチルコリンという化学的信号は再び活動電位という電気的信号に戻る。
⑥このようにして発生した活動電位は横行小管(T管)上にあるジヒドロピリジン受容体:DHPR(電位依存性カルシウムチャネル:Cav1.1=L型1)に伝わる。
⑦活動電位が伝わるとジヒドロピリジン受容体は構造が変化する。
⑧ジヒドロピリジン受容体の構造が変化すると筋小胞体上にあるカルシウムチャネルであるリアノジン受容体Ⅰ型が活性化されてカルシウムイオンが筋小胞体から放出されることになる。
⑨放出されたカルシウムイオンがトロポニンに結合するとトロポミオシンがアクチンから引き離されて、ミオシン頭部がアクチンと結合できるようになる。こうしてアクチンとミオシンは相互作用(滑走)ができるようになり筋収縮が起こる。
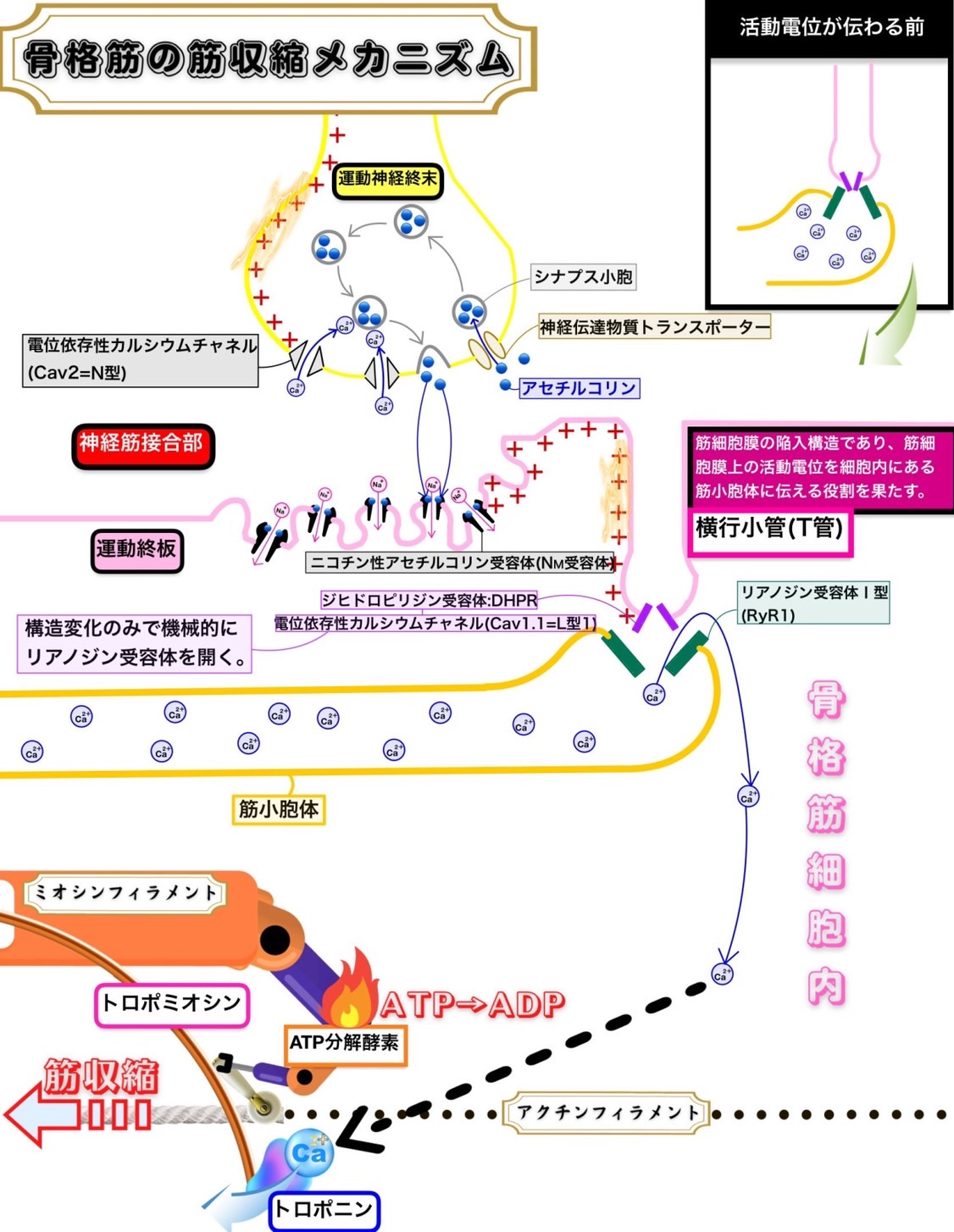
心筋の筋収縮メカニズム
心臓拍動の自動中枢である洞房結節が1回活動電位が発生させると心臓は1回拍動する。
つまり、心筋の収縮は洞房結節が発生させた活動電位が刺激伝導系を通じて伝播していく結果として起こっているのである。
通常、心筋細胞は隣接した細胞に活動電位が発生しない限りは静止膜電位を保ち脱分極することはない。
しかし、洞房結節細胞(房室接合部周辺やHis束の心筋細胞も洞房結節細胞ほどではないが自動能を持つ)は外部からの刺激が無くても内因性に活動電位を発生させて自動的に興奮する自動能を持つ。
また、刺激伝導系の心筋細胞も自動能を持つものの通常は洞房結節細胞に追従する形となっている。刺激伝導系上位の細胞から十分な電位が伝わらない場合は、下位の細胞が自動能を発揮して補充する。
骨格筋が運動神経の指令によって筋収縮を起こしている機構とは異なることに注意する必要がある。
心筋と骨格筋の筋収縮メカニズムの違い

活動電位発生機構の違い
心筋は不随意筋であるのに対して、骨格筋は随意筋である。このため、骨格筋細胞が運動神経の指令によって脱分極を生じて筋収縮を起こしている機構と心筋が収縮する機構は異なる。
心筋細胞は隣接した心筋細胞の活動電位が伝播した結果として脱分極を生じる。
洞房結節細胞が自動能を持っており、刺激伝導系を通じて心臓全体に活動電位を行き渡らせるペースメーカーとして機能している。
また、心筋細胞に活動電位が発生した結果として起こる心筋収縮までの機構も骨格筋細胞のものとは異なる。
活動電位の信号を細胞内に伝える機構の違い
心筋ではジヒドロピリジン受容体:DHPR(電位依存性カルシウムチャネル:Cav1.2=L型2)からのカルシウムイオン流入がカルシウムイオン依存的にリアノジン受容体Ⅱ型を活性化する。これは構造変化のみでリアノジン受容体Ⅰ型を活性化させる骨格筋にあるジヒドロピリジン受容体:DHPR(電位依存性カルシウムチャネル:Cav1.1=L型1)とは作用の仕方が異なる。
つまり、心筋の収縮の際に筋小胞体からカルシウムイオンが放出されるためにはリアノジン受容体Ⅱ型を活性化するためにカルシウムイオンのジヒドロピリジン受容体:DHPR(電位依存性カルシウムチャネル:Cav1.2=L型2)を通した心筋細胞内流入が必要であり、この筋小胞体からのカルシウムイオン放出様式をカルシウムイオン誘発性カルシウムイオン放出という。
一方で、骨格筋の収縮の際は筋小胞体からカルシウムイオンが放出されるためにカルシウムイオンの骨格筋細胞内流入が必要がなく、このカルシウムイオン放出様式を脱分極誘発性カルシウムイオン放出という。ジヒドロピリジン受容体とリアノジン受容体には種類があり、筋収縮に関わる方法がそれぞれ異なるということである。
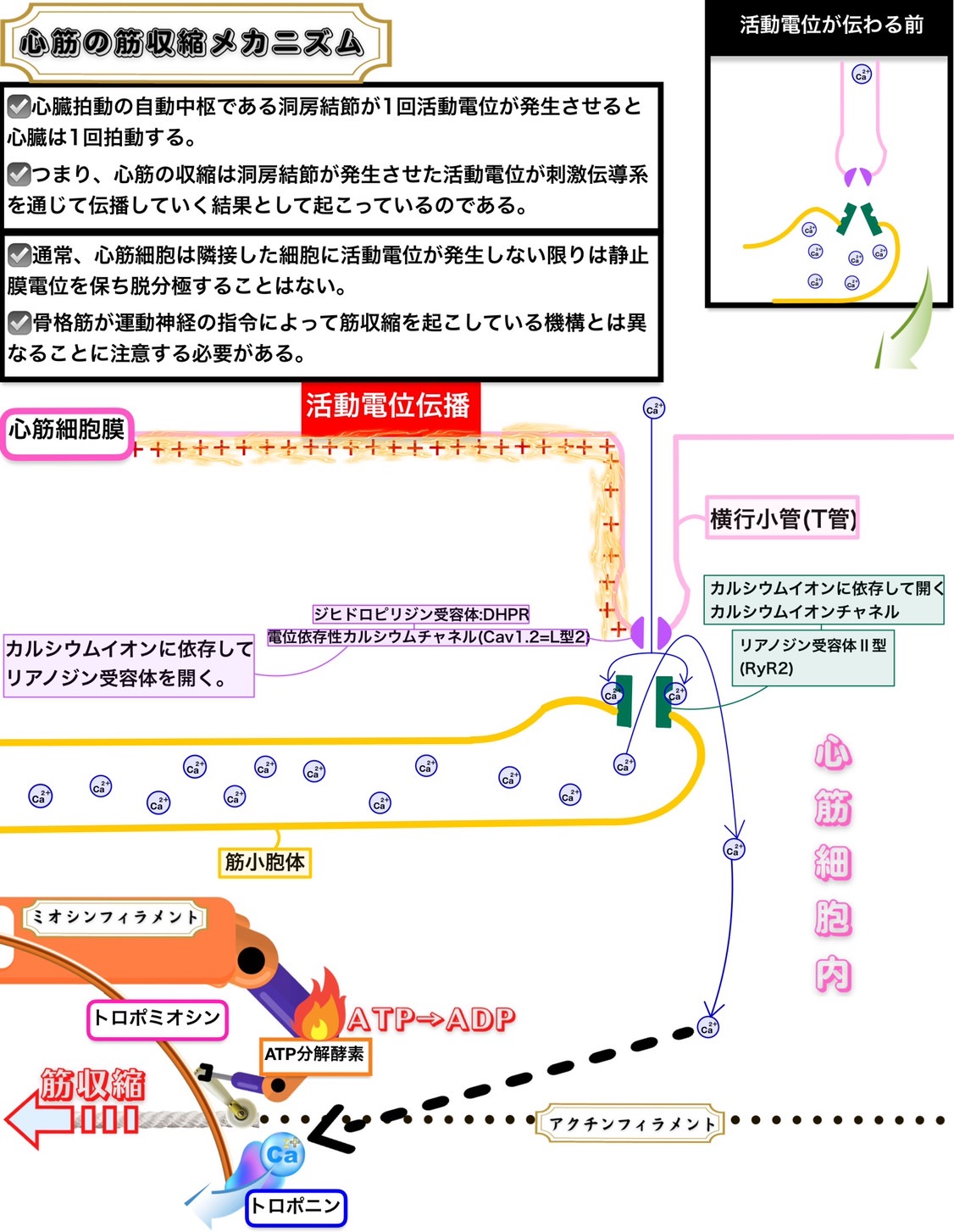
滑り説
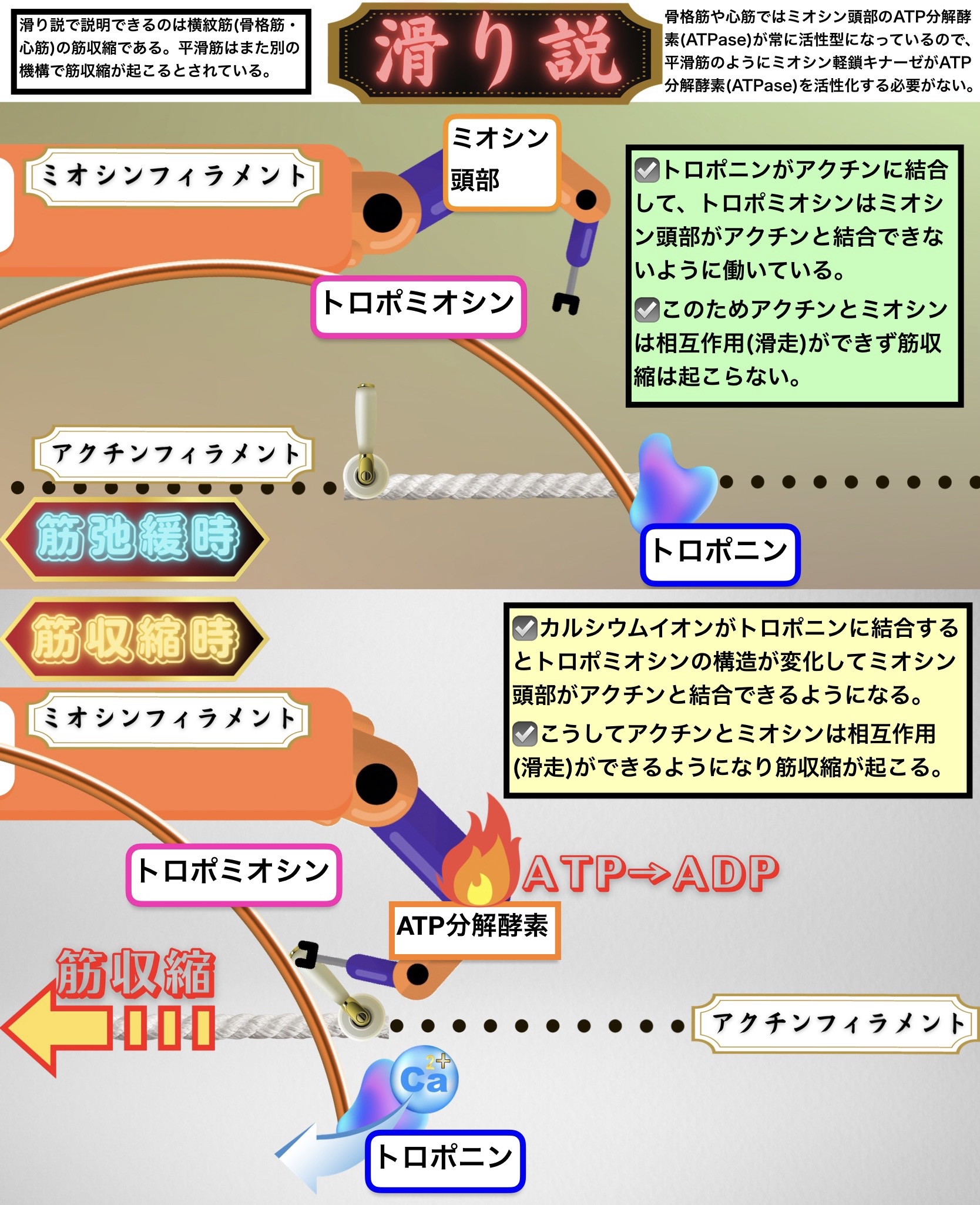
滑り説で説明できるのは横紋筋(骨格筋・心筋)の筋収縮である。平滑筋はまた別の機構で筋収縮が起こるとされている。
筋弛緩時
トロポニンがアクチンに結合して、トロポミオシンはミオシン頭部がアクチンと結合できないように働いている。
このためアクチンとミオシンは相互作用(滑走)ができず筋収縮は起こらない。
筋収縮時
カルシウムイオンがトロポニンに結合するとトロポミオシンの構造が変化してミオシン頭部がアクチンと結合できるようになる。
こうしてアクチンとミオシンは相互作用(滑走)ができるようになり筋収縮が起こる。
血管平滑筋収縮・弛緩のメカニズム
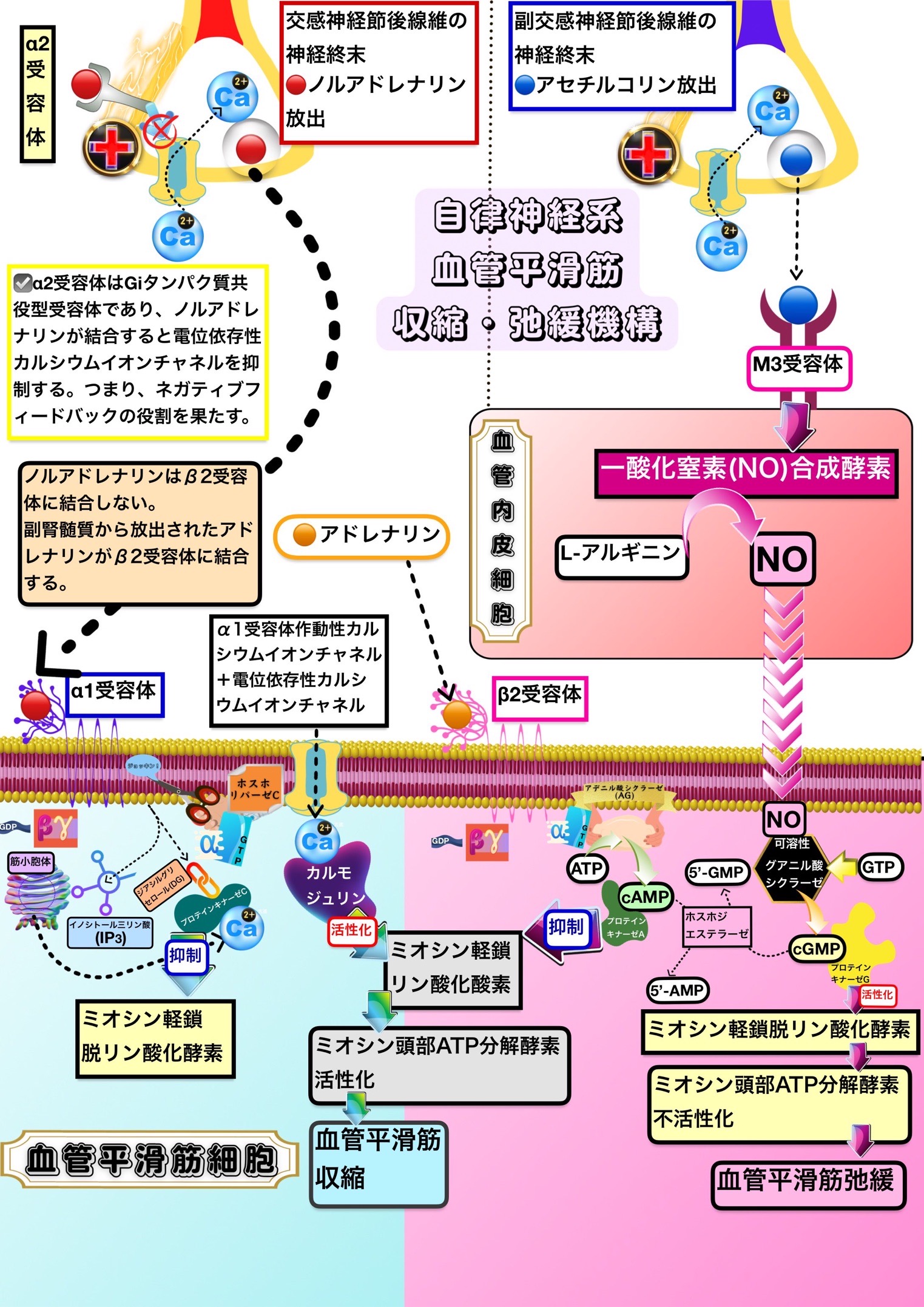
平滑筋の筋収縮は滑り説では説明できない

滑り説で説明できるのは横紋筋(骨格筋・心筋)の筋収縮である。
平滑筋には横行小管が存在しない。また、トロポニンも存在しない。
このため、平滑筋の筋収縮は滑り説では説明することができない。
平滑筋の筋収縮・筋弛緩の鍵を握っているのは「ミオシン頭部ATP分解酵素の活性化・不活性化」である。
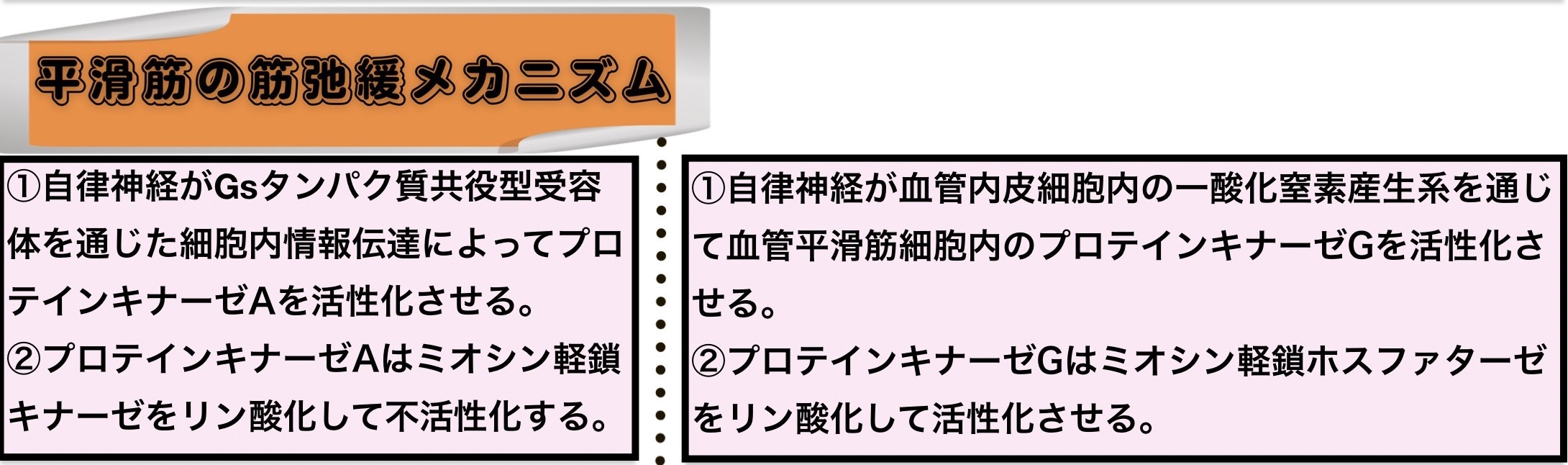
平滑筋を筋収縮させる機構はミオシン頭部ATP分解酵素を活性化させるためにミオシン軽鎖キナーゼ(ミオシン軽鎖リン酸化酵素)を活性化させてミオシン軽鎖ホスファターゼ(ミオシン軽鎖脱リン酸化酵素)を抑制するように働き、平滑筋を筋弛緩させる機構はミオシン頭部ATP分解酵素を不活性化させるためにミオシン軽鎖キナーゼを抑制してミオシン軽鎖ホスファターゼを活性化させるように働く。
骨格筋や心筋ではミオシン頭部のATP分解酵素(ATPase)が常に活性型になっているので、平滑筋のようにミオシン軽鎖キナーゼがATP分解酵素(ATPase)を活性化する必要がない。
平滑筋の筋収縮メカニズム

①機械的な刺激や脱分極によって電位依存性カルシウムイオンチャネルが開いて細胞外から細胞内にカルシウムイオンが流れ込む。また、自律神経が受容体作動性カルシウムイオンチャネルを開いて細胞外から細胞内にカルシウムイオンを流し込んだり、Gqタンパク質共役受容体を通じてIP3誘発性カルシウムイオン遊離を引き起こす。
②平滑筋細胞内でカルシウムイオン濃度が高まるとカルシウムイオンはカルシウムイオン結合タンパク質であるカルモジュリンと結合する。
③カルシウムイオンと結合したカルモジュリンはミオシン軽鎖キナーゼ(ミオシン軽鎖リン酸化酵素)を活性化する。
④また、Gqタンパク質共役受容体を通じて活性化されたプロテインキナーゼCはミオシン軽鎖ホスファターゼ(ミオシン軽鎖脱リン酸化酵素)を抑制することで筋収縮を促進する。
平滑筋の筋弛緩メカニズム

コメント