
骨格筋の筋収縮メカニズム
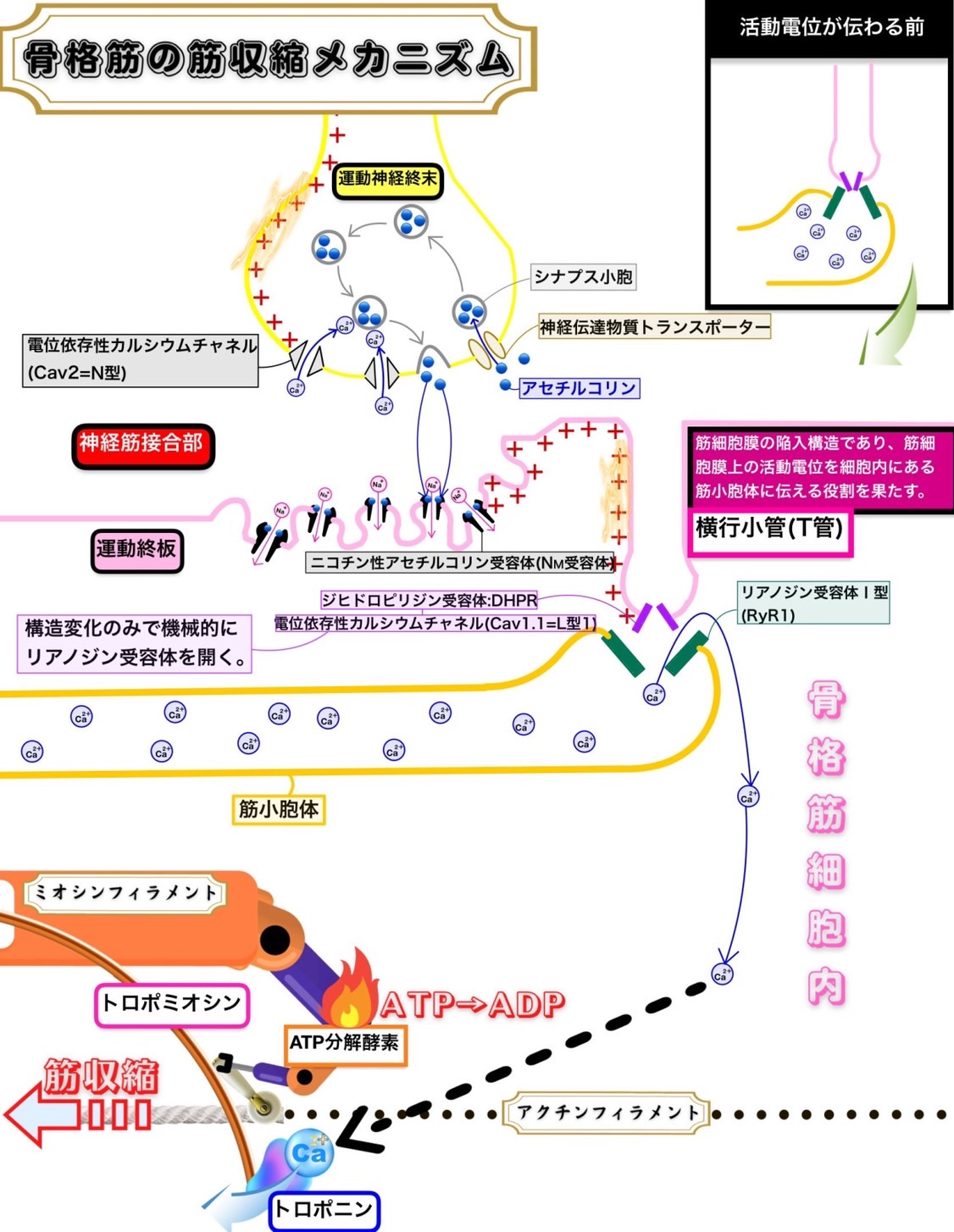

①中枢からの骨格筋収縮の指令は活動電位により電気的信号として運動神経線維を伝わってきて運動神経終末までやってくる。運動神経線維は自律神経のように中枢から支配器の間で神経節でニューロン交代(乗り換え)をすることがないので、節前線維と節後線維という区別がなく1本である。
②活動電位が伝わるとシナプス前膜が脱分極して電位依存性カルシウムイオンチャネルが開く。
③カルシウムイオンが運動神経終末に流入してきて運動神経細胞内のカルシウムイオン濃度が上がると、シナプス小胞がシナプス前膜と膜融合してアセチルコリンをシナプス間隙に開口分泌(exocytosis)する。ここで活動電位という電気的信号がアセチルコリンという化学的信号に変わることになる。
④放出されたアセチルコリンは運動終板上にあるニコチン性アセチルコリン受容体(Nm受容体)に結合する。
⑤ニコチン性アセチルコリン受容体(Nm受容体)にアセチルコリンが結合するとナトリウムイオンが流入して運動終板が脱分極する。ここでアセチルコリンという化学的信号は再び活動電位という電気的信号に戻る。
⑥このようにして発生した活動電位は横行小管(T管)上にあるジヒドロピリジン受容体:DHPR(電位依存性カルシウムチャネル:Cav1.1=L型1)に伝わる。
⑦活動電位が伝わるとジヒドロピリジン受容体は構造が変化する。
⑧ジヒドロピリジン受容体の構造が変化すると筋小胞体上にあるカルシウムチャネルであるリアノジン受容体Ⅰ型が活性化されてカルシウムイオンが筋小胞体から放出されることになる。
⑨放出されたカルシウムイオンがトロポニンに結合するとトロポミオシンがアクチンから引き離されて、ミオシン頭部がアクチンと結合できるようになる。こうしてアクチンとミオシンは相互作用(滑走)ができるようになり筋収縮が起こる。
心筋の筋収縮メカニズム
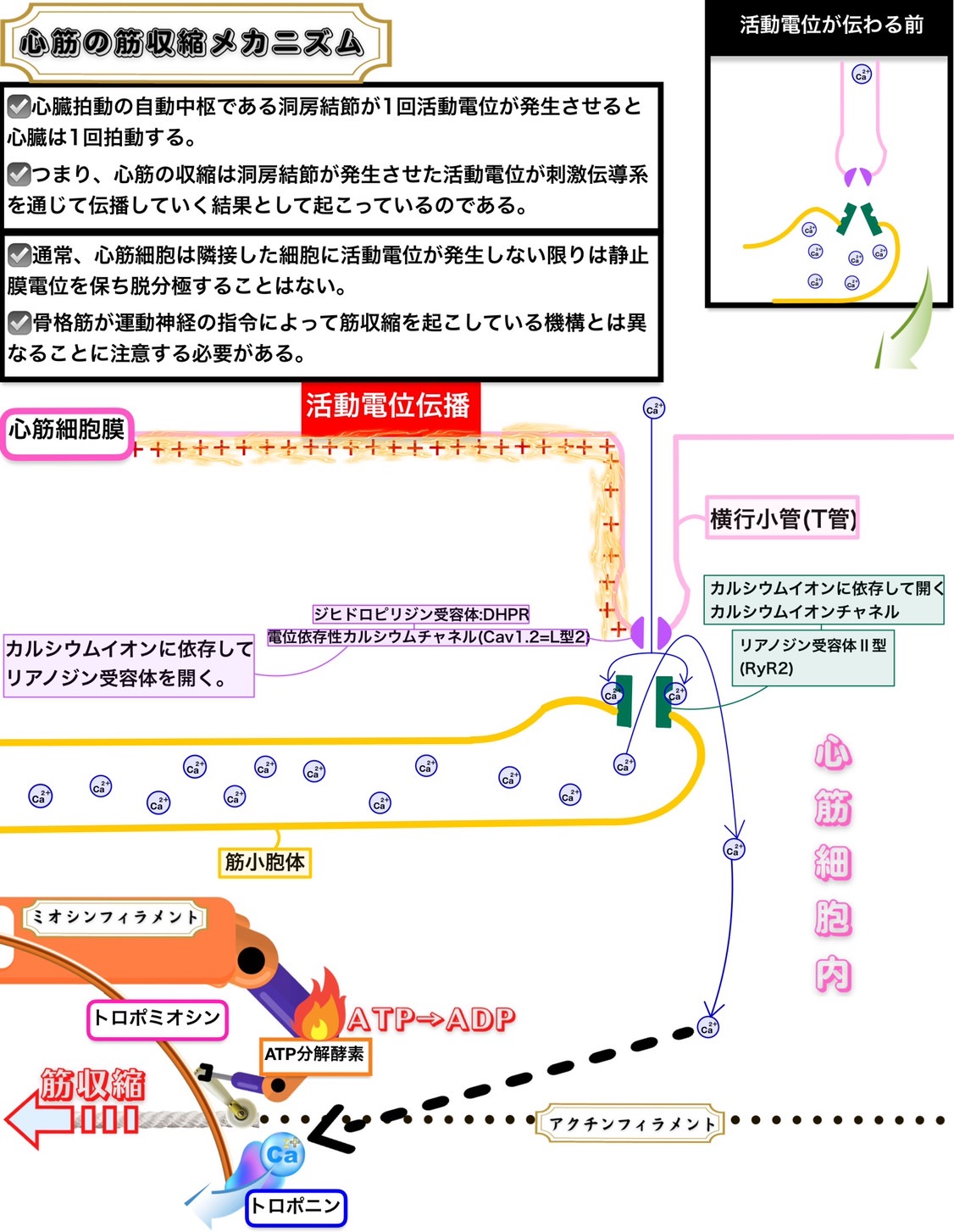
心臓拍動の自動中枢である洞房結節が1回活動電位が発生させると心臓は1回拍動する。
つまり、心筋の収縮は洞房結節が発生させた活動電位が刺激伝導系を通じて伝播していく結果として起こっているのである。
通常、心筋細胞は隣接した細胞に活動電位が発生しない限りは静止膜電位を保ち脱分極することはない。
しかし、洞房結節細胞(房室接合部周辺やHis束の心筋細胞も洞房結節細胞ほどではないが自動能を持つ)は外部からの刺激が無くても内因性に活動電位を発生させて自動的に興奮する自動能を持つ。
また、刺激伝導系の心筋細胞も自動能を持つものの通常は洞房結節細胞に追従する形となっている。刺激伝導系上位の細胞から十分な電位が伝わらない場合は、下位の細胞が自動能を発揮して補充する。
骨格筋が運動神経の指令によって筋収縮を起こしている機構とは異なることに注意する必要がある。
心筋と骨格筋の筋収縮メカニズムの違い

活動電位発生機構の違い
心筋は不随意筋であるのに対して、骨格筋は随意筋である。このため、骨格筋細胞が運動神経の指令によって脱分極を生じて筋収縮を起こしている機構と心筋が収縮する機構は異なる。
心筋細胞は隣接した心筋細胞の活動電位が伝播した結果として脱分極を生じる。
洞房結節細胞が自動能を持っており、刺激伝導系を通じて心臓全体に活動電位を行き渡らせるペースメーカーとして機能している。
また、心筋細胞に活動電位が発生した結果として起こる心筋収縮までの機構も骨格筋細胞のものとは異なる。
活動電位の信号を細胞内に伝える機構の違い
心筋ではジヒドロピリジン受容体:DHPR(電位依存性カルシウムチャネル:Cav1.2=L型2)からのカルシウムイオン流入がカルシウムイオン依存的にリアノジン受容体Ⅱ型を活性化する。これは構造変化のみでリアノジン受容体Ⅰ型を活性化させる骨格筋にあるジヒドロピリジン受容体:DHPR(電位依存性カルシウムチャネル:Cav1.1=L型1)とは作用の仕方が異なる。
つまり、心筋の収縮の際に筋小胞体からカルシウムイオンが放出されるためにはリアノジン受容体Ⅱ型を活性化するためにカルシウムイオンのジヒドロピリジン受容体:DHPR(電位依存性カルシウムチャネル:Cav1.2=L型2)を通した心筋細胞内流入が必要であり、この筋小胞体からのカルシウムイオン放出様式をカルシウムイオン誘発性カルシウムイオン放出という。
一方で、骨格筋の収縮の際は筋小胞体からカルシウムイオンが放出されるためにカルシウムイオンの骨格筋細胞内流入が必要がなく、このカルシウムイオン放出様式を脱分極誘発性カルシウムイオン放出という。ジヒドロピリジン受容体とリアノジン受容体には種類があり、筋収縮に関わる方法がそれぞれ異なるということである。
滑り説
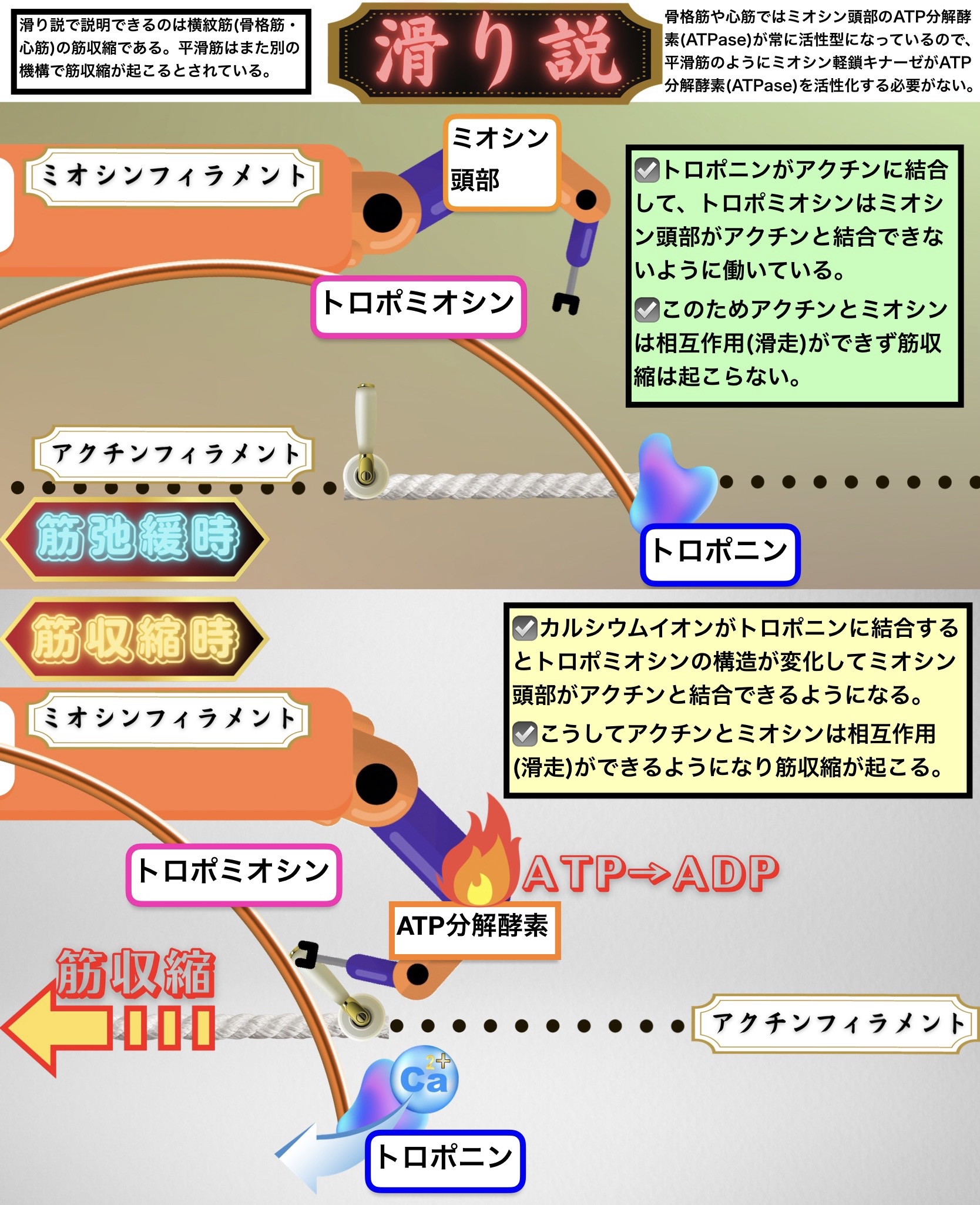
滑り説で説明できるのは横紋筋(骨格筋・心筋)の筋収縮である。平滑筋はまた別の機構で筋収縮が起こるとされている。
筋弛緩時
トロポニンがアクチンに結合して、トロポミオシンはミオシン頭部がアクチンと結合できないように働いている。
このためアクチンとミオシンは相互作用(滑走)ができず筋収縮は起こらない。
筋収縮時
カルシウムイオンがトロポニンに結合するとトロポミオシンの構造が変化してミオシン頭部がアクチンと結合できるようになる。
こうしてアクチンとミオシンは相互作用(滑走)ができるようになり筋収縮が起こる。
血管平滑筋収縮・弛緩のメカニズム
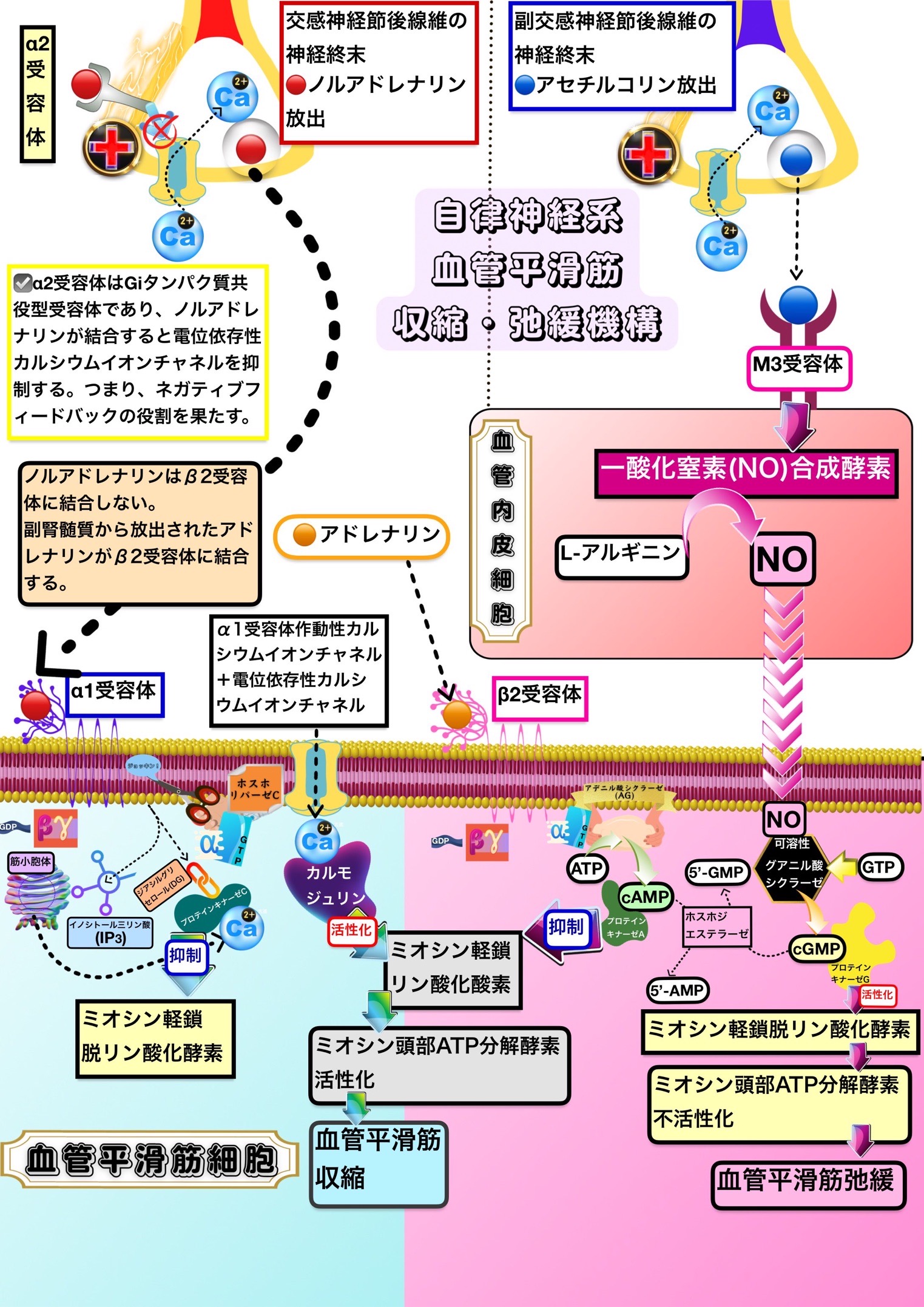
平滑筋の筋収縮は滑り説では説明できない

滑り説で説明できるのは横紋筋(骨格筋・心筋)の筋収縮である。
平滑筋には横行小管が存在しない。また、トロポニンも存在しない。
このため、平滑筋の筋収縮は滑り説では説明することができない。
平滑筋の筋収縮・筋弛緩の鍵を握っているのは「ミオシン頭部ATP分解酵素の活性化・不活性化」である。
平滑筋を筋収縮させる機構はミオシン頭部ATP分解酵素を活性化させるためにミオシン軽鎖キナーゼ(ミオシン軽鎖リン酸化酵素)を活性化させてミオシン軽鎖ホスファターゼ(ミオシン軽鎖脱リン酸化酵素)を抑制するように働き、平滑筋を筋弛緩させる機構はミオシン頭部ATP分解酵素を不活性化させるためにミオシン軽鎖キナーゼを抑制してミオシン軽鎖ホスファターゼを活性化させるように働く。
骨格筋や心筋ではミオシン頭部のATP分解酵素(ATPase)が常に活性型になっているので、平滑筋のようにミオシン軽鎖キナーゼがATP分解酵素(ATPase)を活性化する必要がない。
平滑筋の筋収縮メカニズム

①機械的な刺激や脱分極によって電位依存性カルシウムイオンチャネルが開いて細胞外から細胞内にカルシウムイオンが流れ込む。また、自律神経が受容体作動性カルシウムイオンチャネルを開いて細胞外から細胞内にカルシウムイオンを流し込んだり、Gqタンパク質共役受容体を通じてIP3誘発性カルシウムイオン遊離を引き起こす。
②平滑筋細胞内でカルシウムイオン濃度が高まるとカルシウムイオンはカルシウムイオン結合タンパク質であるカルモジュリンと結合する。
③カルシウムイオンと結合したカルモジュリンはミオシン軽鎖キナーゼ(ミオシン軽鎖リン酸化酵素)を活性化する。
④また、Gqタンパク質共役受容体を通じて活性化されたプロテインキナーゼCはミオシン軽鎖ホスファターゼ(ミオシン軽鎖脱リン酸化酵素)を抑制することで筋収縮を促進する。
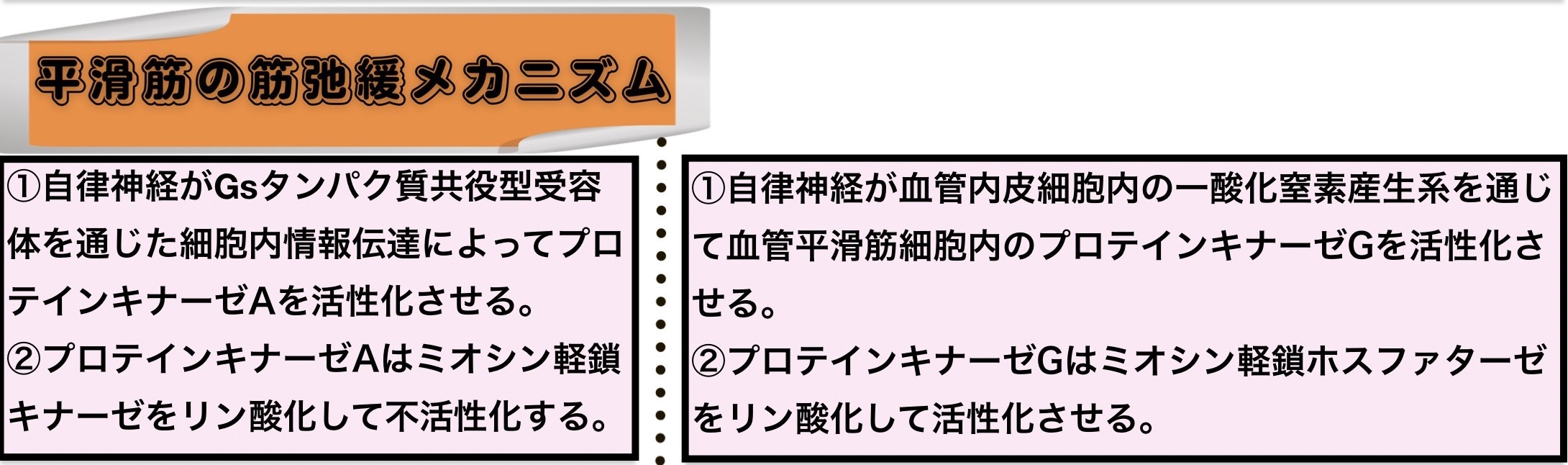
平滑筋の筋弛緩メカニズム

コメント
Very energetic post, I enjoyed that a lot. Will there be a part2?
I like this post, enjoyed this one thank you for posting. «Fear not for the future, weep not for the past.» by Percy Bysshe Shelley.
Having read this I believed it was very informative. I appreciate you taking the time and effort to put this information together. I once again find myself spending way too much time both reading and commenting. But so what, it was still worth it!
Thanks for one as marvelous posting! I quite enjoyed reading it,
This is a good tip especially to those fresh to the blogosphere. Short but very precise info… Appreciate your sharing this one. A must read article.
There is definately a great deal to find out about this subject. I really like all the points you have made. Larina Nels Abrams
This is a great tip particularly to those new to the blogosphere. Simple but very accurate information Appreciate your sharing this one. A must read post!
What’s up, after reading this remarkable paragraph i am as well glad toshare my experience here with friends.
chloroquine malaria hydroxychloroquine and zinc
I think this is a real great blog.Really looking forward to read more. Great.
When I initially commented I clicked the «Notify me when new comments are added» checkbox and now each time a comment is added I get three e-mails with the same comment.Is there any way you can remove me from that service?Bless you!
Great, thanks for sharing this blog.Thanks Again. Want more.
I enjoyed reading this post such an interesting info post. Thanks for providing good info.
A fascinating discussion is worth comment. There’s no doubt that that you should write more about this topic, it might not be a taboo matter but typically people don’t discuss such issues. To the next! Cheers!!
Really nice pattern and great content material , very little else we require : D.
Heya! I just wanted to ask if you ever have any trouble with hackers? My last blog (wordpress) was hacked and I ended up losing many months of hard work due to no back up. Do you have any solutions to prevent hackers?
Great information. Lucky me I found your blog by chance (stumbleupon).I have saved as a favorite for later!
modafinil alternatives – modapls modafinil moa
I have read so many articles or reviews concerning the blogger loversbut this paragraph is truly a nice article, keep it up.
İnstagram: Şifresiz Takipçi Hilesi — Sifresiz-Takipci.com
Nice read, I just passed this onto a friend who was doing a little research on that. And he just bought me lunch as I found it for him smile Thus let me rephrase that: Thank you for lunch!
I have been surfing on-line more than 3 hours lately, but I never discovered any fascinating article like yours. It is lovely value enough for me. In my view, if all webmasters and bloggers made just right content as you probably did, the internet might be much more useful than ever before.
It’s actually a nice and helpful piece of info. I’m glad that you shared this helpful information with us. Please keep us up to date like this. Thanks for sharing.
F*ckin’ tremendous issues here. I am very satisfied to look your post. Thanks so much and i’m taking a look ahead to touch you. Will you kindly drop me a mail?
Howdy just wanted to give you a brief heads up and let you know a few of the images aren’t loading correctly.I’m not sure why but I think its a linking issue.I’ve tried it in two different internet browsers and both show the same outcome.
I’m impressed, I must say. Actually rarely do I encounter a blog that’s both educative and entertaining, and let me inform you, you’ve got hit the nail on the head. Your thought is excellent; the problem is something that not sufficient people are speaking intelligently about. I am very glad that I stumbled throughout this in my seek for something regarding this.
Hello my family member! I wish to say that this post is amazing, nice written and come with approximately all significant infos. I would like to look more posts like this .
460033 395365I would like to see much more posts like this!.. Great blog btw! reis Subscribed.. 531691
Major thanks for the blog post.Really thank you! Awesome.
Thanks-a-mundo for the article post.Much thanks again. Much obliged.
Thanks , I have recently been searching for information approximately this topic for ages and yours is thebest I have discovered till now. But, what about theconclusion? Are you certain in regards to the supply?
When someone writes an piece of writing he/she keeps the idea of a user in his/her mindthat how a user can be aware of it. So that’s why this post is great.Thanks!
Precisely what I was searching for, appreciate it for putting up.
Very good blog article.Thanks Again.
Looking forward to reading more. Great blog article. Great.
apartments for rent in greenville sc rentberry scam ico 30m$ raised ambassador apartments
Awesome blog.Thanks Again. Fantastic.
Thanks for sharing, this is a fantastic article.Really thank you! Awesome.
Thanks for sharing your thoughts about pof. Regards
Whoa! This blog looks exactly like my old one! It’s on a entirely different subject but it has pretty much the same layout and design. Excellent choice of colors!
You could certainly see your skills in the paintings you write. The sector hopes for even more passionate writers such as you who are not afraid to mention how they believe. All the time go after your heart.
The design look great though! Hope you get the problem resolved soon. Cheers
Im obliged for the post.Really looking forward to read more.
I loved your article post. Will read on…
Hi there colleagues, its fantastic post concerning cultureand completely explained, keep it up all the time.
Thank you for your article.
Enjoyed every bit of your blog.Thanks Again. Fantastic.
Awesome blog article.Thanks Again. Fantastic.
It’s an amazing article designed for all the online viewers;they will get advantage from it I am sure.Feel free to surf to my blog post: Semzia Brain Review
Major thanks for the article post.Much thanks again. Fantastic.
Appreciate you sharing, great article. Really Cool.
Thanks a lot for the blog article.Thanks Again. Awesome.
Wow a lot of great knowledge!essay writing tools thesis papers for sale write my thesis
Awesome blog.Really looking forward to read more. Awesome.
I am so grateful for your blog post. Really Great.
Im grateful for the blog post.Really thank you! Much obliged.
Fine way of explaining, and nice article to obtain facts about mypresentation subject, which i am going to present in university.
I really enjoy the article.Much thanks again. Want more.
The quantity to the proper is the stake, and the quantityto the left is the winnings.
online slot games vegas slots online vegas slots online
Asking questions are truly fastidious thing if you are not understanding something fully, howeverthis article provides nice understanding even.
When compared with several other suppliers Rolex encompasses a huge global recognition.
I value the article. Really Great.
Asking questions are genuinely nice thing if you are not understanding something fully, except this article presents pleasant understanding yet.
This is a really good tip especially to those fresh to the blogosphere. Brief but very accurate infoÖ Many thanks for sharing this one. A must read article!
Very informative blog post. Really Cool.
Aw, this was a really good post. Finding the time and actual effort to produce a really good articleÖ but what can I sayÖ I put things off a lot and never seem to get anything done.
I really enjoy the article post.Really thank you! Much obliged.
Aw, this was an extremely good post. Spending some time and actual effort to create a good articleÖ but what can I sayÖ I procrastinate a whole lot and don’t seem to get anything done.
665378 128859I see which you are employing WordPress on your blog, wordpress may be the very best. :~- 588565
Hello, yes this piece of writing is really good and I have learned lot of things from it on the topic of blogging.thanks.
I want to to thank you for this good read!! I absolutely enjoyed every little bit of it. I’ve got you saved as a favorite to check out new things you post…
college essay writing help – essay helper online help with essays
Hydroquinone: Uses, Side Effects, Risks, And More
What Is Hydroquinone and What Are Its Uses?
Hydroquinone, also known as 1,4-dihydroxybenzene, is a natural skin lightener derived
from aromatic plants like bearberry or green tea. It is commonly
used to reduce pigmentation and improve skin texture.
Hydroquinone works by inhibiting the production of melanin, the pigment responsible for dark spots
and uneven skin tones. While it has been used for centuries
in traditional medicine, its application must be done carefully due
to potential side effects.
How Hydroquinone Works
Hydroquinone works by blocking the enzyme tyrosinase, which is responsible for melanin production. By inhibiting
this enzyme, it helps fade hyperpigmentation and reduces dark spots over time.
Its effectiveness is often compared to other skin lighteners, with results usually visible within a few months of consistent use.
Skin Conditions That Benefit From Its Use
Hydroquinone is particularly effective for treating several skin conditions:
Melasma: A condition characterized by dark patches on the face,
often caused by hormonal changes. Hydroquinone helps
reduce pigmentation and lighten these areas.
Eczema: While not a cure, hydroquinone can help alleviate the redness and inflammation associated
with eczema when applied to affected areas.
Acne Scars: By inhibiting melanin production, hydroquinone can help fade
the dark scars left by acne.
Psoriasis: Hydroquinone may help reduce the appearance
of psoriatic scales and patches when used as part of
a broader treatment plan.
Age Spots: One of the most common uses, hydroquinone effectively lightens age-related hyperpigmentation on the face and hands.
Getting an Expert Opinion
Before using hydroquinone, it is crucial to consult with
a dermatologist. They can assess your skin condition, determine if hydroquinone is appropriate for you, and provide guidance on safe application methods.
Is a Skin-Lightening Cream Safe for All Skin Types?
While hydroquinone is generally considered safe when used as directed, it can cause irritation or burns in some individuals.
Those with sensitive skin, rosacea, or eczema may want to avoid using hydroquinone-based products.
How to Use It
Apply hydroquinone cream as recommended by your dermatologist,
typically once daily in the morning and evening.
Always follow the dosage instructions on the product label and avoid applying it in excess or without guidance
from a healthcare professional.
What Happens When You Stop
If you discontinue using hydroquinone, you may notice a
recurrence of dark spots as melanin production resumes.
Consistent use is key to maintaining results, so consider
long-term strategies for skin health with your doctor’s advice.
Potential Risks and Side Effects
Common side effects include mild irritation or burns, especially in sensitive areas like the face.
Overuse can lead to peeling or dryness, so it is important to adhere to recommended
application frequencies. Long-term use should be
discussed with a dermatologist to assess potential health risks.
Natural Alternatives to Hydroquinone
If you are considering alternatives, natural options like antioxidants (Vitamins A and
C) and plant-based acids can offer similar benefits without the risks associated with hydroquinone.
These ingredients work synergistically to reduce pigmentation and promote skin health.
Antioxidants Such as Vitamins A and C
Vitamin C is particularly effective at brightening skin tone
and protecting against oxidative stress, while Vitamin B3 (niacinamide) helps reduce melanin production and
improves skin elasticity. Together, these antioxidants can provide natural alternatives to hydroquinone.
Plant-Based Acids
Plant-based acids like bearberry extract contain hydroquinone,
which can help lighten skin pigmentation naturally.
However, excessive use may lead to irritation, so moderation is key when using these
products.
Vitamin B3
Vitamin B3 (niacinamide) is another natural alternative
that helps reduce hyperpigmentation and improve skin texture.
It works by inhibiting melanin transfer in the skin, leading to a brighter complexion over time.
Summary
Hydroquinone is an effective treatment for certain skin conditions, but its use must be approached with caution due to
potential side effects and risks. Consulting with a dermatologist is essential
before starting any new skincare regimen. Natural
alternatives like antioxidants and plant-based
acids can offer similar benefits without the associated health risks.
A Word From Verywell
“Hydroquinone can be an effective treatment for certain skin conditions, but it should only be used under medical supervision,”
says Dr. Jane Kim, a dermatologist at Verywell.
“The potential for irritation and long-term health effects necessitates careful consideration and professional guidance.”
Frequently Asked Questions
Q: Is hydroquinone safe for all skin types?
A: Not necessarily. It may cause irritation in individuals with sensitive skin conditions like rosacea or eczema.
Q: How long does it take to see results from hydroquinone?
A: Results typically appear within a few months of consistent use, but this
can vary depending on the individual and the severity of their condition.
Q: Are there natural alternatives to hydroquinone?
A: Yes, antioxidants like Vitamins A and C, as well as plant-based acids,
can provide similar benefits without the associated risks of hydroquinone.
Q: Can hydroquinone be used on the face?
A: While it is effective for facial conditions like melasma, it should only be used under
medical supervision to avoid irritation or burns.
My blog what do steroids do to women’s bodies
Thanks-a-mundo for the article.Thanks Again. Really Cool.
I think this is a real great article.Really looking forward to read more. Great.
Great, thanks for sharing this article.Much thanks again. Really Great.
Say, you got a nice article post.Really looking forward to read more. Really Cool.
Very good blog article. Much obliged.
Muchos Gracias for your blog. Great.
Pretty! This was an incredibly wonderful article. Many thanks for providing this info.
Hi there! Do you know if they make any plugins to assist with SEO?I’m trying to get my blog to rank for some targetedkeywords but I’m not seeing very good gains. If you know of any please share.Many thanks!
I value the blog article.Really thank you! Awesome.
If you are searching for the ultimate pest control company, look no more. These guys are awesome! The best out there.
Enjoyed every bit of your blog article. Want more.
I really like reading a post that can make people think. Also, many thanks for allowing me to comment.
Great, thanks for sharing this article post.Really looking forward to read more. Will read on…
In fact, a systematic graphical representation is done by professional designers who have creative bent
It is really a nice and helpful piece of info. I’m happy that you simply shared this helpful info with us.Please keep us up to date like this. Thanks for sharing.
Im obliged for the blog post.Thanks Again.
Looking forward to reading more. Great blog post.Much thanks again. Will read on
Wow, great article post. Really Cool.
This is my first time visit at here and i am in fact pleassant to read all at single place.
Thanks in favor of sharing such a good thinking, paragraph is pleasant, thats why ihave read it completely
แทงบอลออนไลน์บอกเลยครับผมว่าคอบอลไม่ควรพลาดเพราะว่าคุณสามารถทำการพนันได้อย่างง่ายๆโดยไม่ต้องเดินทางไปโต๊ะบอลเหมือนสมัยก่อนเพียงแต่คุณเข้ามาที่ UFABET แล้วลงทะเบียนเป็นสมาชิกคุณก็สามารถพนันได้เลยในทันที ฝากถอนไม่มีขั้นต่ำใช้ระบบอัตโนมัติ สะดวกปลอดภัย
I really enjoy the blog article.Really looking forward to read more. Great.
modafinil online provigil medication modafinil uses
I wanted to thank you for this excellent read!! I definitely enjoyed every bit of it. I’ve got you saved as a favorite to look at new stuff you postÖ
Really enjoyed this blog.Really thank you! Fantastic.
Hello there! This is my first visit to your blog! We are a team of volunteers and starting a new project in a community in the same niche. Your blog provided us valuable information to work on. You have done a extraordinary job!
Great, thanks for sharing this article post.Thanks Again. Fantastic.
Hey There. I found your blog using msn. This is an extremely well written article. I’ll be sure to bookmark it and come back to read more of your useful info. Thanks for the post. I’ll definitely comeback.
Howdy! This post could not be written any better! Reading through this post reminds me of my good old room mate! He always kept chatting about this. I will forward this post to him. Fairly certain he will have a good read. Thanks for sharing!
Muchos Gracias for your blog.Really looking forward to read more. Really Cool.
I think this is a real great blog post.Really looking forward to read more. Really Great.
Very informative article.Thanks Again.
Very nice post. I just stumbled upon your blog and wanted to say that I have really enjoyed surfing around your blog posts. In any case I?ll be subscribing to your rss feed and I hope you write again soon!
It is in reality a nice and useful piece of info. I’m glad that you simply shared this helpful information with us. Please stay us up to date like this. Thank you for sharing.
Wow, great post.Much thanks again. Keep writing.
Thank you for your blog post. Keep writing.
Hello my friend! I wish to say that this post is awesome, nice written and include approximately all vital infos. I’d like to see more posts like this.
Very neat blog.Really thank you! Great.
Pretty! This was an incredibly wonderful article. Thank you for supplyingthis information.
For you to know your chances of winning, you require to know firsthow lottery works.
Really informative post.Really thank you! Really Cool.
It is in point of fact a nice and useful piece of information. I am glad that you shared this useful info with us.Please stay us informed like this. Thanks for sharing.
Hello, everything is going fine here and ofcourseevery one is sharing facts, that’s truly good, keep up writing.
Hello, yup this article is actually pleasant and Ihave learned lot of things from it concerning blogging.thanks.
ivermectin manufacturer ivermectin solubility
colleges with no essays required google essay writer essay writers geek
Sweet blog! I found it while browsing on Yahoo News. Do you
have any tips on how to get listed in Yahoo
News? I’ve been trying for a while but I never seem to
get there! Many thanks
https://telegra.ph/Top-kazino-11-14-2
https://t.me/s/officials_casino_1win
https://t.me/mcasino_martin/677
https://t.me/mcasino_martin/754
https://t.me/officials_pokerdom/3158
Hey! I know this is kinda off topic however , I’d figured I’d ask.
Would you be interested in trading links or maybe guest authoring a blog post
or vice-versa? My website discusses a lot of the same topics as yours
and I think we could greatly benefit from each other. If you are interested feel free to send me an e-mail.
I look forward to hearing from you! Excellent blog by the way!
My coder is trying to convince me to move to .net from PHP.
I have always disliked the idea because of the expenses.
But he’s tryiong none the less. I’ve been using Movable-type on various websites for about a year
and am anxious about switching to another platform.
I have heard great things about blogengine.net. Is there a way I can transfer all my wordpress
content into it? Any kind of help would be really appreciated!
Great, thanks for sharing this article.Much thanks again. Fantastic.
Say, you got a nice blog article.Really thank you! Really Great.
https://t.me/s/iGaming_live/4873
Very good article post.Much thanks again. Will read on…
Great, thanks for sharing this article post.Really thank you! Great.
I was curious if you ever thought of changing the structure of your site?
Its very well written; I love what youve got to say. But maybe you could a little more in the way
of content so people could connect with it better.
Youve got an awful lot of text for only having one or two
pictures. Maybe you could space it out better?
http://images.google.ki/url?q=https://t.me/officials_7k/383